
When we consider movement in humans (or animals for that matter), we must start out with the understanding that there is no such thing as a prescription for perfect movement.
It is sometimes tempting to think we could hone the body to some type of perfection where it would move perfectly—and we can be easily seduced into thinking of the body in a mechanistic way, like a car whose performance can be finely tuned.
However, with a car the performance is task-specific. A Formula One car can travel with exhilarating speed and agility around the Silverstone Grand Prix circuit but would struggle in the streets of London, and fare even worse on a farmer’s field – while other vehicles are built for these purposes.
The human organism is obviously different from a machine, but one particular difference is worth spelling out loud and clear: It wouldn’t matter how many times you drove a Ferrari around a field, its performance would not improve; in fact, it would very likely get worse due to the damage it would suffer from the inappropriate terrain. A human being, on the other hand, would improve his or her performance at a different or unfamiliar task, and continue to progress until a satisfactory level of competence had been achieved.
If we do a similar job or activity for most of our lives, we will become both mentally and physically skilled at it. At this point the car analogy may look like it fits a little better; we are really good at the thing we do regularly and much less good at the things we do infrequently. The car is built for a purpose, human beings become fit for purpose.
The Mechanistic Model
Most of us will be familiar with the model of muscles shown below, with one or two muscles acting as agonist and antagonist over a joint to effect movement at the joint. It is certainly the model I was introduced to during my osteopathic studies. If we open almost any anatomy book we will see descriptions like this:
Biceps Brachi
Origin: coracoid process of scapula
Insertion: tuberosity of radius
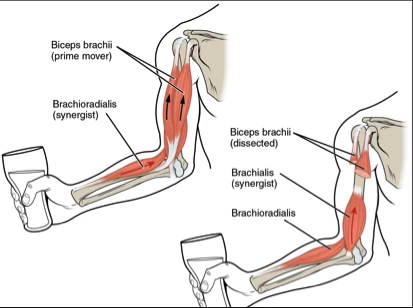 Action: Flexes the elbow joint moving the forearm towards the humerus
Action: Flexes the elbow joint moving the forearm towards the humerus
Innervation: C5, C6
Every muscle in the body can be described in such a way. There is some truth in this explanation of the muscular system, but it’s a half-truth that gets us to picture the body in a segmented way. It’s a Newtonian model of movement and a whole science of exercise and manual therapy has been built around it.
It is appealing and fairly easy to grasp as a concept. If someone has a problem with an activity and you analyze it from this perspective, it would make sense to target the muscles you felt were failing, by strengthening the weak muscles and lengthening the tight ones so that the human machine will balance itself and function better. This would be a little like getting the tracking sorted out in the front wheels of your car to reduce wear and tear on the tires.
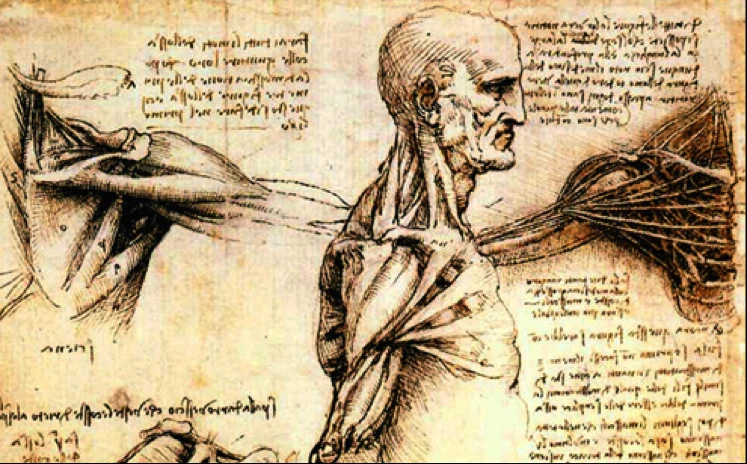
Since nearly all anatomy has been studied through dissection, as these wonderful drawings by Leonardo da Vinci show, it was not unreasonable to ascribe certain movements to the muscles they found under the skin. Leonardo preceded Isaac Newton by more than a hundred years, but much of the anatomical research that went on beyond Leonardo was done to the backdrop of the mechanistic discoveries of Newton and those that followed him. The mechanistic human machine was a powerful idea.
These ideas were given further impetus in the latter part of the 19th century, when areas of the brain were found to control certain muscular actions. There was much debate about how parts of the cerebral cortex controlled movement, but the argument seemed to be settled when, in 1937, Wilder Penfield, after painstaking research, published his map of the motor and sensory cortex.
Perhaps because Penfield took the step to publish it using illustrations, it stayed in the imagination of those who study the nervous system in a way that words never could. Penfield had a frustration though: however carefully he stimulated the motor cortex, he frequently didn’t get the response he expected; a muscle he didn’t expect to move would move. He put this down to his equipment, which he felt was not sufficiently delicate to stimulate the discrete areas he intended, and thus there was resultant overlapping of stimulation.
However, in 1971, Hiroshi Asanuma, with much more sophisticated equipment, tried to map the motor cortex even more accurately. Using micro stimulation techniques with ultra-fine wire electrodes, he intended to stimulate one neuron at a time, hoping to find an absolute one-to-one relationship between motor cortex areas and individual muscles. Asanuma used the finest wire electrodes possible and the smallest voltage possible to get a detectable result and reduce the possibility of overlap. He also used the shortest duration possible to avoid damage to the tissues being stimulated. Even then – frustratingly for the experimenter – a small amount of overlap still occurred; here was another example of mechanistic thinking at work.
What Asanuma, and Penfield before him, had set out to prove was that if you stimulated a neuron in the motor cortex, the impulse would travel down the nerve and excite the muscle at the other end. The analogy of throwing a light switch and the bulb lighting up is obvious and appealing.
All this research was thrown into question in the late 1990s. As often happens, an accidental observation in an animal-testing laboratory revealed a very different possibility about the way the motor cortex is laid out. A researcher had been attempting to research the frontal eye field, where the electrode stimulation lasted for half a second rather than the much briefer five milliseconds used to study the motor cortex.
He inadvertently missed the frontal eye field and got the electrode into the primary motor cortex. When the half-second stimulation was applied, the monkey extended its arm forwards and shaped its fingers as if to reach for something. A whole movement pattern unfolded.
These observations led to a new line of research where stimulation was extended somewhat to see what happened. The results were astonishing and led to an overturning of the discrete muscle theory.
What researchers Michael Graziano, Tirin Moore and Charlotte Taylor discovered was that the motor cortex was mapped into behavioral repertoires. The stimulation of one neuron didn’t lead to a downward pulse along the line of the nerve to the muscle; rather it spread out to recruit other neurons causing a whole group of muscles to be recruited into a recognizable movement pattern. What previous researchers had noted as a discrete muscle twitch was, in fact, the tip of a movement iceberg. It was the pursuit of a preconceived idea—i.e direct muscle-to-neuron relationship—that stopped them seeing the overlaps they were getting for what they really were: the start of a behavioral movement pattern.
More on this subject from YogaUOnline and Peter Blackaby – Movement Transformations Part 2: Understanding the Nature of Functional Movement.
Reprinted with permission from www.intelligentyoga.co.uk

Peter Blackaby has 30-plus years’experience as a yoga teacher and trainer, and is also trained as an Osteopath. Peter started practicing yoga seriously in 1978 as a student of the Iyengar System and after six years undertook the two-year teacher training program, qualifying in 1986. Currently, he teaches functional anatomy on the London yoga teacher-training course and has input to two other local courses. He is regularly invited to teach throughout England, Wales, and Scotland.
The project he is presently involved with is a two-year course for teachers called Grounded Yoga. The aim is to help students understand how yoga can bring productive change to the body, the mind, and the breath and improve our sense of relationship to the environment in which we find ourselves. Find more about Peter Blackaby at www.intelligentyoga.co.uk
